Genome Editing is the ability to make specific changes at targeted genomic sites and is of fundamental importance to researchers in biology and medicine. Two genome editing technologies have emerged recently that exploit bacterial systems for plant pathogenesis or adaptive immunity:
Both the technologies use endonucleases that initiate double-strand breaks (DSBs) at virtually any genomic target sequence. While both technologies are popular, the decision to choose one technology over the other is not always clear. Therefore, depending on several factors such as Specificity, Target site selection, efficiency and ease of design, one can choose the best suited technology.
What is CRISPR?
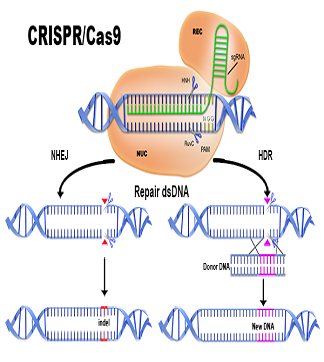
CRISPR stands for Clustered Regularly Interspaced Short Palindromic Repeats. CRISPR sequences were originally identified in the Escherichia coli (E. coli) genome, and were found to function as part of an RNA-based adaptive immune system to target and destroy genetic parasites at the DNA level.
CRISPR-associated protein (Cas) is an endonuclease that cuts foreign DNA, allowing integration into the host genome. Cleavage only occurs when there is a protospacer adjacent motif (PAM) around the targeted sequence of the invading DNA, ensuring highly accurate targeting.
Researchers studying CRISPR have adapted it for use as a tool for genetic modification of the target host genome. CRISPR/Cas9 has recently become a popular genome editing tool, due to its simplicity and versatility.
CRISPR Services
CRISPR/Cas9 genome editing has become widely used due to its simplicity and versatility, and the CRISPR technology has been adapted for diverse applications aside from genome editing. As a leader in gene synthesis and genome editing, and through our partnership with Feng Zhang’s laboratory at the Broad Institute of MIT and Harvard*, GenScript offers validated CRISPR products, services and resources to help you harness the power of CRISPR genome editing for your research
What is TALEN?
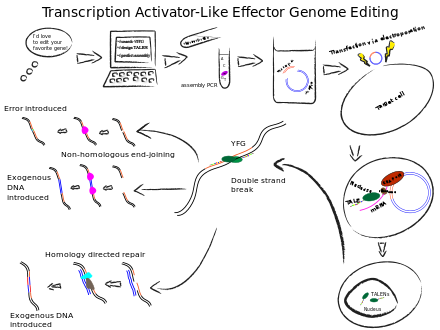
TALENs operate on almost the same principle as ZFNs. TALENs are fusions of transcription activator-like (TAL) proteins and a FokI nuclease. TAL proteins are composed of 33-34 amino acid repeating motifs with two variable positions that have a strong recognition for specific nucleotides (Deng et al., 2014). By assembling arrays of these TALs and fusing them to a FokI nuclease, specific cutting of the genome can be achieved. When two TALENs bind and meet, the FokI domains induce a double-strand break which can inactivate a gene, or can be used to insert DNA of interest (Cermak et al., 2011). TALENs are more specific than ZFNs, the monomers don’t have the cross-reactivity problem that plagued ZFN researchers. This simplicity has facilitated automation of TALEN construction (Reyon et al., 2012). On the downside, since a TAL is needed for each nucleotide, TALENs are larger and somewhat harder to deliver than ZFNs.